Store energy and save energy
Many devices have been developed to harvest energy from walking or running, but their use often comes at cost to the wearer in the form of increased metabolic demand. Shepertycky et al. designed a device that can harvest mechanical energy from a natural walking gait and convert it to useable electrical energy while also reducing the metabolic energy consumption of the user (see the Perspective by Riemer et al.). The key to achieving “something from nothing” comes from designing the device to use muscle-centric control of the knee exoskeleton resistance to reduce active muscle force during the late part of the leg swing cycle.
Science, aba9947, this issue p. 957; see also abh4007, p. 909
Abstract
Evolutionary pressures have led humans to walk in a highly efficient manner that conserves energy, making it difficult for exoskeletons to reduce the metabolic cost of walking. Despite the challenge, some exoskeletons have managed to lessen the metabolic expenditure of walking, either by adding or storing and returning energy. We show that the use of an exoskeleton that strategically removes kinetic energy during the swing period of the gait cycle reduces the metabolic cost of walking by 2.5 ± 0.8% for healthy male users while converting the removed energy into 0.25 ± 0.02 watts of electrical power. By comparing two loading profiles, we demonstrate that the timing and magnitude of energy removal are vital for successful metabolic cost reduction.
Humans are exceptional walkers, with our gait continuously adapting over evolutionary (1) and developmental time scales (2). In the absence of frictional losses, walking on level ground at a constant speed theoretically requires no energy input (3), as passive mechanisms permit the exchange of kinetic and potential energy (4, 5). In reality, however, walking is metabolically expensive. Walking requires more energy than any other activity of daily living (6, 7). During walking, humans expend metabolic energy, in part, to restore dissipated kinetic energy (8).
Exoskeletons have been developed to assist the lower-limb muscles in restoring dissipated kinetic energy to improve walking efficiency. Active (powered) exoskeletons use actuators, such as electric motors, to perform a portion of the positive joint work that is naturally done by concentric muscle contractions (9, 10). These devices essentially add energy to the human-device system to supplement the metabolic energy required for walking (fig. S1). Passive (unpowered) exoskeletons use elastic mechanisms to aid users’ muscle-tendon units in storing and transferring energy from one gait phase to another. By assisting energy recycling, these devices decrease the force that muscle-tendon units must hold during isometric contractions (11).
Although existing devices assist users by adding or recycling energy, is it possible to improve walking efficiency by purely removing energy? This would require a different design approach: Instead of assisting muscles in concentric or isometric contractions, devices would need to assist muscles in producing force during eccentric contractions. The strategy of removing energy is similar to those employed by some biomechanical energy harvesters optimized to produce electrical power (12–14). Unfortunately, these devices have not been able to provide their users with a metabolic benefit over carrying their own weight, let alone an advantage over normal walking [(12–14); also see supplementary materials]. Because the metabolic cost of walking can be explained primarily by the cost of actively generating muscle force (15), we hypothesized that we could optimize these devices for metabolic cost reduction by designing them with the aim of decreasing muscle activation instead of producing electrical power.
Assisting muscles in producing force during eccentric contractions is challenging. Substantial negative joint work during walking results from the passive deformation of soft tissues (8), such as ligaments. These tissues do not require metabolic energy and so cannot be metabolically assisted. Furthermore, some soft tissues store and return energy that would otherwise be dissipated (5, 8); thus, disrupting these energy return mechanisms could lead to a substantial energetic penalty because the body must compensate through concentric contractions. Because eccentric muscle contractions are already metabolically efficient compared with concentric contractions (16), assisting muscles in eccentric contractions should yield only a small energetic advantage. Therefore, assisting muscles in eccentric contractions may require the exoskeleton to apply a load more accurately and precisely, regarding both magnitude and timing, than is required for concentric contraction and powered exoskeletons (9, 10, 17).
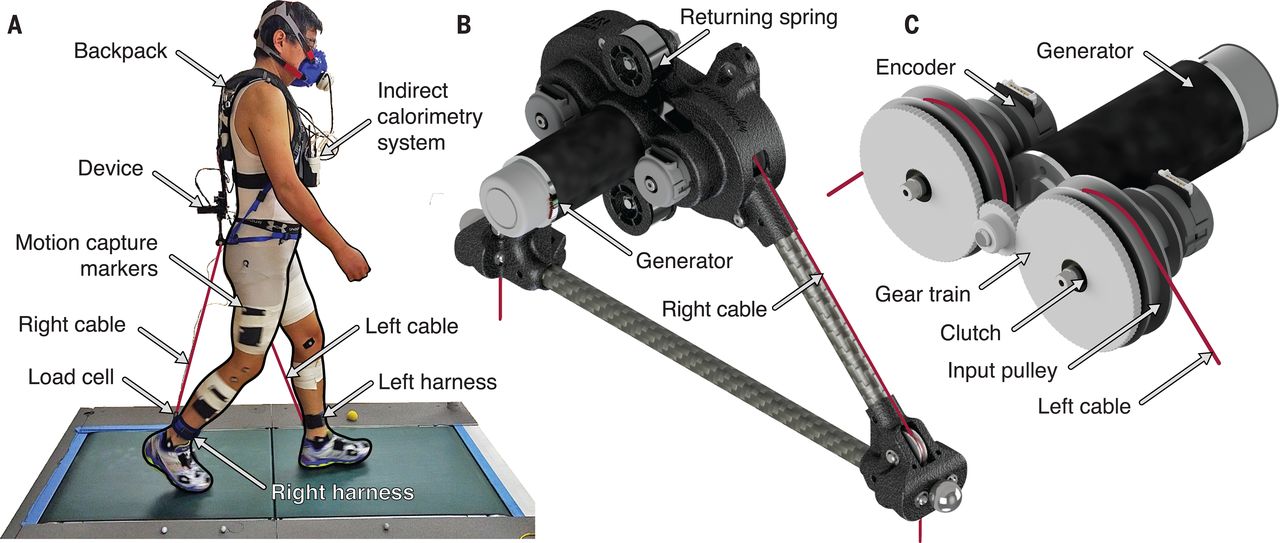
We built a lightweight, backpack-mounted exoskeleton (1.059 kg; Fig. 1) to assist the hamstrings in producing force by applying a knee flexion moment during the terminal swing phase of the gait cycle. This device consists of cables that are attached by harnesses to the user’s lower legs and extend upward to connect to input pulleys. As the user’s knee extends forward during the swing period, the swinging leg’s input cable is unspooled and drives the input pulley. These pulleys work in combination with a gear train and roller clutches to convert the cables’ linear movements into rotational motion while simultaneously amplifying and integrating the cables’ motions before engaging a single generator. During stance, the clutches decouple the input pulleys from the gear train, allowing the cables to be gathered back onto the input pulleys by returning springs. The clutches also enable the device to apply mechanical load only during the swing period, leaving the lower limbs unencumbered by the exoskeleton during the stance period. The back electromotive force of the generator applies the mechanical load to the input cables and, in turn, to the user (further described in the supplementary materials, movies S1 and S2, and data S3).
(A) A participant walking on the split-belt force-sensing treadmill while wearing the exoskeleton and measurement equipment. (B) Outer view of the device. (C) View of the device’s internal components.
” data-icon-position=”” data-hide-link-title=”0″>
(A) A participant walking on the split-belt force-sensing treadmill while wearing the exoskeleton and measurement equipment. (B) Outer view of the device. (C) View of the device’s internal components.
Biomechanical energy harvesters traditionally apply a mechanical loading profile onto their users that is proportional to the motion input into the device (12–14). This technique is simple to implement because it requires no active control, but it may sacrifice user assistance. In this study, we implemented an active control system that enabled our device to apply a controllable mechanical load by regulating the generator’s back electromotive force. Exoskeletons have successfully reduced the metabolic energy requirement of walking by using assistive loading profiles that mimic either natural joint moment or power profiles (18–21). However, these profiles do not account for the contribution of passive elements that cannot be assisted. To specifically target the hamstrings and avoid interfering with natural energy return mechanisms, we implemented a profile that resembles the muscles’ contribution to the net knee moment during the terminal swing phase. We derived this muscle-centric profile by subtracting the estimated contribution of passive elements, based on results of Whittington et al. (22), from the net knee moment profile. We then scaled the magnitude of the resultant profile to the user’s body mass. The control system and loading profiles are described in the supplementary materials, figs. S2 and S3, and data S4.
We conducted treadmill-walking experiments with 10 healthy male participants to determine the metabolic and biomechanical effects of the muscle-centric profile compared with both a traditional harvesting profile and normal walking. Minimal changes in both knee joint angles and net joint moments indicate that neither loading profile hindered normal knee function (Fig. 2, A to C), which can cause deviations from normal walking such as a stiff knee gait (14). The lack of effect on normal knee function was further confirmed by a lack of changes in spatial-temporal parameters such as stride length and ground contact time. The average angles, moments, and powers for the ankle, knee, and hip joints are presented in fig. S4 and data S5 and S6.
(A) Knee joint angles over a gait cycle for the three walking activities: normal walking (gray), traditional profile (blue), and muscle-centric profile (red). (B) Average stride lengths. (C) Net knee moment over a gait cycle normalized to body mass. (D) Average estimated negative muscle moment contribution during the swing period. (E) Moment applied by the device about the user’s knee over a gait cycle. (F) Average mechanical power removed by the device. (G) Percentage of the negative muscle impulse applied by the device during the swing period. (H) Hamstring muscle activity over a gait cycle. (I) Average hamstring muscle activity for an entire gait cycle and specific gait phases (right groupings). *P < 0.05, ***P < 0.001. Data are presented as means ± SEMs. Flex., flexion; Ext., extension; Int., initial; resp., response.
” data-icon-position=”” data-hide-link-title=”0″>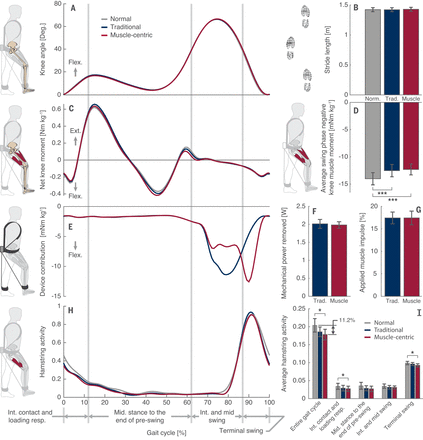
(A) Knee joint angles over a gait cycle for the three walking activities: normal walking (gray), traditional profile (blue), and muscle-centric profile (red). (B) Average stride lengths. (C) Net knee moment over a gait cycle normalized to body mass. (D) Average estimated negative muscle moment contribution during the swing period. (E) Moment applied by the device about the user’s knee over a gait cycle. (F) Average mechanical power removed by the device. (G) Percentage of the negative muscle impulse applied by the device during the swing period. (H) Hamstring muscle activity over a gait cycle. (I) Average hamstring muscle activity for an entire gait cycle and specific gait phases (right groupings). *P < 0.05, ***P < 0.001. Data are presented as means ± SEMs. Flex., flexion; Ext., extension; Int., initial; resp., response.
One difference between the two loading profiles (Fig. 2E) was that the moment applied about the user’s knee by the traditional profile peaked during mid-swing (80.1 ± 0.5% gait cycle, mean ± SEM), whereas that applied by the muscle-centric profile peaked during terminal swing (90.7 ± 0.2% gait cycle). Both loading profiles significantly reduced the estimated average muscle contribution to the negative net knee moment during the swing period [n = 10 participants; repeated-measures analysis of variance (ANOVA), F2,18 = 17.806, P < 0.001, ηp2 = 0.664], with the traditional profile reducing it by 11.0 ± 1.8% [least significant difference (LSD) post hoc analysis: P < 0.001] and the muscle-centric profile reducing it by 12.0 ± 2.0% (P = 0.001; Fig. 2D).
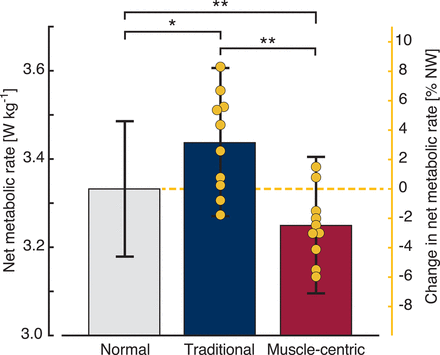
Although both profiles removed a similar amount of mechanical power (traditional: 2.01 ± 0.12 W, muscle-centric: 1.98 ± 0.09 W; Fig. 2F) and applied a similar percentage of the muscles’ angular impulses (traditional: 17.4 ± 1.3%, muscle-centric: 17.4 ± 1.5%; Fig. 2G), the muscle-centric profile reduced the energy requirement of walking by 2.5 ± 0.8% [n = 10; one-way repeated-measures ANOVA; F2,18 = 12.714, P < 0.001, ηp2 = 0.586; LSD post hoc analysis: mean diff. (MD) = −0.082 ± 0.024 W kg−1, P = 0.008, 95% CI (−0.137, −0.027)], whereas the traditional profile increased the energy requirement by 3.1 ± 1.1% [MD = 0.106 ± 0.039 W kg−1, P = 0.024, 95% CI (0.018, 0.195)]. The difference between profiles was 0.19 ± 0.05 W kg−1 [MD = 0.188 ± 0.046 W kg−1, P = 0.003, 95% CI (0.085, 0.291); Fig. 3 and table S1].
The average net metabolic rate for the three walking activities (gray, normal walking; blue, traditional profile; red, muscle-centric profile) is normalized to the participant’s body mass (mean ± SEM). The left vertical axis represents the net metabolic rate (means and error bars; watts per kilogram). The right vertical axis represents the metabolic rate as a percent change from normal walking (NW; means and scatter). As such, no scatter is included for the normal walking condition. Net metabolic rate significantly differed between walking conditions (*P < 0.05, **P < 0.01). Each yellow circle represents one participant.
” data-icon-position=”” data-hide-link-title=”0″>